
Breeding & Genetics
the Tricolor Cockatiel
plus halfsiders and other anomalies
Note: the sex chromosomes of birds actually function according to the ZW sex-determination system and this is the terminology used by scientific sources. It's customary in the pet-bird community to use XY terminology because it's easier to remember, and this article follows that custom.
1. What is tricolor?a. Minor terminology issue
2. Halfsiders - not quite the same thing
3. Tricolor observations
4. Mixed DNA as a possible genetic cause
a. Chimeras
b. Gynandromorphs
c. Genetic mosaics
5. Mosaic or chimera?
6. Can these anomalies be a family trait?
7. Other possible causes
8. What is the third color in a tricolor cockatiel?
9. Technical details - migration of embryonic pigment cells
a. More speculation on the pied gene and multiple-gene interactions
b. Speculation on color changes over time
c. Other information on mosaicism that probably isn't relevant
10. Conclusion
11. Photo gallery
A tricolor cockatiel has three colors in its plumage: grey, yellow/white (the orange cheek is lumped into this category), and a third color that typically looks like cinnamon. This color pattern looks rather simple, but the reason for it is a deep, murky mystery with no easy answers. Most cases of abnormal coloration are probably caused by a health problem or diet deficiency, but there are rare cases that appear to be caused by a genetic malfunction. These apparent genetic anomalies are the subject of this article. A photo gallery of tricolor cockatiels and other coloration oddities is presented at the end of the article.
Minor terminology issue. Tricolor cockatiels are sometimes called Schimmel (with a variety of different spellings). "Schimmel" is a German word referring to a dapple or roan horse (one that has mottled or mixed colors on its coat). German breeder Gunter Wulf used the term to describe two of his cockatiels that had unusual plumage, and Terry Martin's cockatiel mutations book published pictures of both birds and described them as Schimmel. However the plumage of these birds is anomalous in different ways. One is clearly a tricolor while the other has a mottled/grizzled appearance that does not look like tricolor. Both can be seen in the photo gallery as Bird #1 and Bird #17.
The word Schimmel is mainly used in Germany to describe a mottled/grizzled look not large patches of solid color, so it's not an ideal label for tricolor cockatiels. But Terry Martin is the leading expert on parrot mutations and terminology, and since he used the word Schimmel to describe a tricolor it's likely that this terminology will persist.
*******************
(Back to Top)
Halfsiders - not quite the same thing
In general it appears that the tricolor phenomenon is somewhat different from the halfsider phenomenon. But it's likely that the underlying genetic cause(s) are somewhat similar and much more information is available on halfsiders than on tricolor, so a discussion of the halfsider phenomenon is warranted.
The word halfsider describes a specific type of unusual physical appearance but doesn't indicate what made the bird look like that. The “classic” halfsider has bilateral asymmetry, meaning different colors on each side that are vertically divided more or less equally right down the middle of the bird. Some sources use the word halfsider to describe any bird with different coloring on different parts of the body that is not explained by ordinary genetic inheritance, but this tends to be confusing if the bird does not have an obvious 'half and half' division in its coloring. Many other color distribution patterns are possible, including quartersiders who only have a partial 'split down the middle' appearance, and birds who have different types of plumage on different parts of the body that are distributed regularly or irregularly with no visible relationship to the centerline of the body.
This article will use the word halfsider to refer to birds that have a vertical 'split down the middle' appearance to some degree. Birds 11-14 in the photo gallery are clear examples of the halfsider phenomenon, and the other birds in the picture gallery are not halfsiders as defined here; only Bird #11 can reasonably be described as both a halfsider and a tricolor. All the other birds in the gallery are one or the other or something else entirely.
There are several recorded examples of halfsiders successfully breeding but they produce normal-colored young. They breed true to whichever "side" the sperm or ovum came from. A family line of halfsiders can not be developed because the halfsider coloring is not inheritable. (Olszewski, Genetics-Psittacine)
The halfsider phenomenon has been observed in budgies (Twinzy, Houdini, budgie 3, budgie 4) and a variety of other pet parrot species including lovebirds (pic 1, pic 2, pic 3, pic 4), IRNs (pic 1, pic 2, pic 3), and eclectus (pic). A 1935 study by Crew and Lamy investigated 17 cases of bicolorism in budgies, 16 of them halfsiders and all of them involving blue and green coloring.
Page 92-93 of Terry Martin's parrot mutations book talks about halfsiders, saying "Genetically these birds are invariably Normal split blue. It is currently believed that they occur due to a genetic error as an embryo. This error deletes the Normal gene from one half of the bird, thereby allowing the Blue gene to express itself in that side of the bird."
Parrots have two types of pigment in the body. Melanin is a black/grey/brown pigment that can be used in combination with feather structure to produce the appearance of blue. Psittacin is a different pigment which produces yellow and red colors, and when yellow psittacin is added to blue coloring it creates the appearance of green. In essence Terry Martin's book said the halfsider phenomenon removes the psittacin pigment from part of the body. In a bird that is normally green and yellow, the part of the body with psittacin will look green and yellow and the part without psittacin will look blue and white. In species like cockatiels where the normal color is NOT green, removing the yellow will have a different visual effect. The halfsider coloring of Birds 13 and 14 in the photo gallery obviously involves the deletion of psittacin pigment, since one side of the body has normal yellow/orange coloring and the other does not.
But in any case, the principle that Martin stated is only partly true. The coloring of most halfsiders does involve the deletion of yellow psittacin from part of the body, but there are other halfsiders where the color difference involves melanin not psittacin, including Bird #11 in the photo gallery. The tricolor phenomenon in cockatiels is a difference in melanin coloring.
The amount of yellow coloring in a bird is controlled by the genes at the blue locus (see the article on Allelic Mutations for a full explanation of the blue locus). Color oddities that involve melanin instead of psittacin are obviously the result of something that happened to a different gene.
*******************
(Back to Top)
The photo gallery indicates that tricolor cockatiels seem to fall into two distinct categories: (1) birds with a recognizable, fairly consistent pattern where the medium tone occurs primarily on the upper wings with a fairly equal distribution on both sides, and normal (or occasionally pied) feathers in the center of the back. Birds 1-7 and are examples of this, and will be called specific-pattern birds in this article; and (2) birds with unique markings that don’t follow any specific pattern and will be called random-pattern birds. It's difficult to classify Bird #8, since his pattern has changed over time; one picture looks fairly similar to the specific pattern and the other looks random. It's probably most accurate to call him a random pattern bird. Birds 9-10 have random patterns that appear to be genetically caused. Birds 15-16 have random patterns that may or may not have a genetic cause.
Birds 11-14 are halfsiders, but only Bird 11 can reasonably be described as tricolor. Birds 15-17 have plumage oddities that can't be described as either halfsider or tricolor, and won't be discussed further in this article.
It seems possible that the specific-pattern tricolor birds are all the result of the same genetic phenomenon, while the random-pattern birds may be the result of different phenomena, which may or may not be related to each other and may or may not be genetic.
Many of the specific-pattern birds are whiteface, and most of the random-pattern birds are not whiteface. Whiteface is an alternate term for the blue gene, which is frequently involved in the halfsider phenomenon in parrots. But there is no obvious relationship between the blue/whiteface gene and the tricolor phenomenon.
All the specific-pattern birds and some of the random-pattern birds are pied, so the tricolor phenomenon may be related to the pied gene somehow. The pied gene affects the migration of melanin-producing cells in the embryo, and it appears that some of these melanin-producing cells or the DNA controlling them are "different" in tricolor birds.
Some but not all of the tricolor birds have the cinnamon gene. It isn't known how the cinnamon-like color is produced on birds that don't have the cinnamon gene. There are a variety of rare mutations that produce lighter and/or browner tones in the feathers, but none of the birds shown here are known to have these genes.
When the pearl mutation is present on a tricolor bird, the pearling doesn’t look normal. General observations of other oddities seem to indicate that the pearl gene is somewhat prone to disruption.
Tricolor birds seem to be generally capable of breeding successfully, but there are no verified cases where a tricolor parent transmitted the tricolor markings to their offspring. However there have been some observations of tricolor and other oddities within the same family.
A post on the Genetics-Psittacine board here describes an example where the cinnamon sister of a tricolor male produced a tricolor baby; the uncle’s color pattern is not described but it sounds like the baby is a random-pattern bird. Two examples are not enough to conclude that random-pattern tricolor might be more reproducible than standard-pattern tricolor, but it’s a possibility. Other sources have stated that some family lines in birds are prone to producing halfsiders, chimeras, and/or genetic mosaics (discussed later).
*******************
(Back to Top)
Mixed DNA as a possible genetic cause
The prevailing opinion is that unusual mixed color patterns are caused by the presence of two or more genotypes (aka two cell lines) in the body. This terminology is clunky and inconvenient, so I'll invent my own term and call it "mixed DNA". Normally, the body is supposed to have the exact same DNA in every cell. But in some situations a bird (or other animal) shows the colors for separate mutations on different parts of its body because it does NOT have the exact same DNA in every cell; instead there are some areas where the DNA is different. There are several ways that this can happen.
(1) With a chimera, two different zygotes fused into one individual during early embryonic development, and the popular belief is that this is where halfsiders come from. The phenomenon is the result of a developmental accident that is the opposite of identical twins - instead of one individual dividing into two, two individuals merge into one.
However, it appears that many if not all "split down the middle" halfsiders are actually genetic mosaics (discussed below) rather than chimeras. It's said that roughly half the mammals on earth are a mosaic to some degree while chimeras are much rarer (except for a technicality involving twin cattle). (Colorado State)
Tetragametic chimeras (the technical term for two zygotes that have blended into one individual) may or may not have visible physical abnormalities. Most chimeras look normal but may have areas of “otherness” throughout the body, where a whole organ has a different DNA type than other areas. There may be some visible signs of chimerism, for example having eyes that are not the same color. The visibility of the different individuals depends on how differentiated the embryos were at the time they merged, and they merge most readily at the earliest stages of development. (Lam, The Tech)
There have been at least two cases where genetic testing concluded that human women were not the mother of their children. The women looked normal so no one knew they were chimeras, and the part of their DNA that was tested did not come from the same "half" that the children did. Once the chimerism was discovered and the DNA from the correct "side" was tested, they were proved to indeed be the mother. (Project Muse)
Chimerism is not inheritable. The parent has mixed DNA but each individual cell in the parent’s body contains just one DNA type, not both. Therefore only one of these DNA types will go into each ovum or sperm.
I can not find any clear evidence on whether or not a chimera can have the "split down the middle" halfsider appearance; some sources say yes and others say no, and none of them give an explanation for their answer. I don't know of any cases where both sides were DNA tested to give a definite answer. The sources describing a halfsider as a tetragametic chimera seem to have assumed that the individual was a chimera based on appearance. Genetic testing is necessary to determine whether the DNA from two different individuals is present or if this is one individual with a genetic difference in part of the body.
(2) Gynandromorphs, aka intersex or hermaphrodite, occur when sex chromosome abnormalities cause the individual to have both male and female characteristics, and the female aspect dominates in some parts of the body while the male aspect dominates in others. With birds, this can cause interesting effects if the bird has one copy of a sex-linked recessive gene. The gene will be expressed in the 'female' area of the body but not in the 'male' area.
There are various causes of gynandromorphy, including (1) a chimera where two zygotes of different sex have merged, or (2) a genetic mosaic (explained in the next section) where a cell divides abnormally during early embryonic development, resulting in some cells with extra sex chromosomes; depending on the exact mix of sex chromosomes in the local cells, some parts of the body follow the XX pattern and others follow XY. An abnormal number of sex chromosomes in a cell could theoretically be inheritable, but most gynandromorphs are sterile so it's basically a moot point.
At least one online source has stated that gynandromorphs are created in chickens when one ovum is fertilized by two sperm (National Geographic). This could certainly cause some interesting DNA effects if it was true, but reliable-looking sources say that this event (polyspermy) is lethal and does not result in viable young (Developmental Biology Interactive). Multiple sperm can penetrate the ovum in birds, but only one will fertilize the ovum and the others will soon disappear. Even if more than one sperm did fertilize the ovum somehow, it couldn't cause gynandromorphy. In birds all sperm carry the X chromosome and do not influence the sex of the zygote; it's the hen who carries the Y chromosome and determines the sex of the offspring. If it was possible to get a live bird from an ovum that was fertilized by two different sperm, the bird would be the same sex on both "sides". Science Mag talks about a study on gynandromorph chickens that has apparently been misunderstood.
(3) A genetic mosaic occurs when cells emerge within an individual with a different DNA pattern than what was originally inherited from the parents. At some point (usually during embryonic development, but it can occur later in life) there is damage, abnormal cell division, or some other spontaneous change to the DNA in one cell, and it continues to proliferate with this changed DNA. All the cells that descend from the changed cell contain this "different" DNA and behave in accordance with its instructions, while the rest of the body's cells carry the original DNA and follow its instructions. The earlier the DNA change occurs during embryonic development, the more numerous and widespread the changed cells will be. A DNA change that occurs later in life may involve a very small area. (Wikipedia, Genetics-Psittacine)
Genetic mosaics are non-inheritable, for the same reason that chimera color patterns are non-inheritable. Each individual cell in the parent’s body contains just one DNA type, so only one DNA type will go into each ovum or sperm.
*******************
(Back to Top)
I’m inclined to think that mosaicism is more likely to be involved in the tricolor phenomenon than chimerism or gynandromorphy, but there’s not enough information to rule anything out. It’s possible that some cases of tricolor might involve one type of mixed DNA while other cases are caused by a different type of mixed DNA.
Mosaics are much more common than chimeras, and it's said that they can have a "split down the middle" halfsider appearance if the first cell division in the zygote is abnormal. If the abnormal division occurs later it can result in smaller patches of the body looking different than the rest. There's more information on the internet about gynandromorphs with this appearance than there is about halfsider mosaics, but the same principle that applies to abnormal divisions of sex chromosomes should also apply to the autosomal chromosomes. (The Tech, Dalton State) The 1935 study of halfsider budgies by Crew and Lamy called the coloring a mosaic and assumed that abnormal division of an autosomal chromosome was involved, but this shouldn't be taken too seriously; at the time the technology didn't exist that would let them examine the DNA that closely.
Most tricolor cockatiels have patches of unusual color not a full-blown halfsider appearance. This suggests that if a mosaic is involved, the DNA change occurred sometime after the embryo's first cell division. But the color differences are extensive enough that the change was probably not too long after this point.
An article by Cryberg argues that halfsider pigeons are chimeras not mosaics. I am not convinced that this is correct however; the reasoning presents an educated opinion not a statement of fact, and the evidence for it isn't all that solid. Unlike most other sources that say chimeras are rare, he says they are quite easy to produce. It's true that they're easy to produce in the laboratory but this uses artificial processes that would not occur in nature (New Republic).
*******************
(Back to Top)
Can these anomalies be a family trait?
Cryberg made an interesting statement about odd coloring occurring in families: "A number of people have reported lines of birds that tended to produce such chimeras regularly". There is a similar statement on page 93 of Terry Martin's parrot mutations book: "whilst certain family lines are predisposed to producing halfsiders, it is impossible to establish them as an inherited mutation". This testimony from two different sources dealing with very different types of birds indicates that some avian family lines might be prone to genetic accidents that result in mixed DNA. But the reason for this is a mystery.
It’s possible that the answer in some cases might be “none of the above”, and some factor other than mixed DNA is responsible for the third color. All the mixed-DNA traits are non-inheritable so there may be some other explanation for the cases where tricolor seems to run in the family. The variability of tricolor patterns suggests that different examples may have different causes, and some of them might be more inheritable than others. It should be noted that the more reliable reports of multiple family members with odd plumage involve random-pattern birds, and it's more difficult to form a hypothesis about the reasons for the coloring in these birds. In some cases the coloring might be a side effect of something else that's going on in the body, such as a biochemical imbalance or an unusually high need for a particular nutrient. These conditions could be hereditary with varying levels of expression.
Unusual coloration may be caused by metabolic disorders and other illnesses, or by injury to the skin or growing feather (Olszewski). Abnormal feather coloring can also be caused by certain nutritional deficiencies. These factors are the mostly likely cause for most cases of abnormally colored feathers, and normal coloring will usually be restored if the underlying problem is corrected. It's expected that mixed DNA will rarely be the cause when abnormal coloring is present, but there are some cases where it appears to be genetic.
As a possible explanation for the halfsider phenomenon, Pilkington suggested that physical injury to delicate embryonic tissues on one side of the neural crest could result in the subsequent daughter cells all having the same defect; in effect, a type of physical scarring proliferates across half the body. This seems unlikely however unless DNA damage is involved.
*******************
(Back to Top)
What is the third color in a tricolor cockatiel?
It looks like the tricolor phenomenon in cockatiels mostly involves a sex-linked gene, cinnamon. Mosaicism could easily explain the phenomenon in a split-cinnamon male, where the normal side of the gene pair was damaged and the cinnamon gene was left in sole control of melanin coloration in the local area, similar to the way that an unpaired cinnamon gene produces visual cinnamon color in a hen.
But what about a tricolor bird who is genetically a visual cinnamon male, a male with no cinnamon genes, or any hen whether she has the cinnamon gene or not? Some speculation from me, based on no evidence at all: in a visual cinnamon male, maybe one of the cinnamon genes breaks in a way that restores some of the normal function or otherwise prevents the undamaged cinnamon gene from taking sole control, so the feather reverts to the default normal grey coloring. Similarly, the sole cinnamon gene of a hen might be changed so the feather reverts to normal grey.
For birds with no cinnamon genes, maybe there is a change in some other gene which results in a cinnamon-looking feather that is not true cinnamon.
If it isn’t cinnamon in some cases, then what is it? The pied gene normally removes all melanin from a pied feather. Is it possible to have an incompletely pied feather, where only some of the melanin is removed and this reduced amount of melanin is spread evenly throughout the feather? Probably not. The pied mutation interferes with the migration of melanoblasts in the embryo. Pied feathers are the result of not having melanin-producing cells in the area, and there's no ability to put melanin in these feathers. There may be cases where there are some melanocytes in the area but not enough to color the whole feather. But it's most likely that this causes grizzled feathers (which are not uncommon in pied cockatiels), not an even distribution of diluted melanin coloring.
There might be some other kind of dilution effect going on, with less melanin being produced in these areas. Or perhaps the amount of melanin is normal, but it's not forming properly so the color is "off". There are known mutations that have similar effects so it's conceivable that the DNA could break in a way that produces this type of coloration in a limited area. The clues still point toward mosaicism or a non-genetic cause.
Susanne Russo asked about the patch of cinnamon feathers on Bird #8 on the Genetics-Psittacine board. The response was “It could be theorised that a local gene defect in that patch of skin caused damage to the cinnamon locus and therefore that patch was unable to convert brown into normal black eumelanin.” The term “genetic mosaic” wasn’t used, but is apparently what was meant, suggesting that a bird with normal grey coloring had localized damage that prevented the melanin pigment from being processed normally.
*******************
(Back to Top)
Technical details - Migration of embryonic pigment cells
Melanoblasts (precursors to melanin-producing cells) arise from the dorsal (top/back) side of the neural crest (the precursor to the brain and spinal cord) and migrate laterally towards the ventral (front/bottom) side of the body where they meet more or less along the midline. This migration pattern explains how the halfsider 'split down the middle' appearance is created when each side of the neural crest is genetically programmed for a different color scheme. Melanoblasts do not arise uniformly along the length of the neural tube; in general, a large pool is present in the cervical region and significantly fewer in the trunk. With piebald/pied mutations, melanoblasts form and begin to disperse but do not migrate very far, so that areas close to the point of origin are more likely to receive melanoblasts than areas that are further away. In birds, pigment cells enter the migratory pathway at the level of the forelimb (wing), which is probably the reason that the upper wing is more likely to retain melanin coloring than any other part of the body. The migration path to the upper wing is shorter than the path to many other parts of the body, so the melanoblasts have a better chance of reaching their destination. (Parichy et al, Osawa, Genetics-Psittacine)
The mechanics of psittacin production and distribution are unknown. But based on the existence of halfsider birds where one side has psittacin pigmentation and the other side does not, it appears that the migration of psittacin-producing cells in the embryo is similar to that of melanin-producing cells.
In humans and a few other mammals, a phenomenon called “Blaschko’s lines” has been observed. This is a pattern of lines on the skin which is invisible in most individuals but may become a visible pattern of stripes or markings in chimeras/mosaics or people with certain skin diseases. It’s believed that these lines show the migration pattern of embryonic cells as the individual developed. These markings tend to be fairly symmetrical. The pattern isn't limited to 'tiger stripes' and there are a variety of other patterns that can occur including large patches of color (Medical Grapevine). Blaschko’s lines have not been observed in birds and are not directly related to the tricolor phenomenon; however it does demonstrate that a DNA change in one cell during an early stage of development may be spread across a wide area of the body following the migration pattern of embryonic cells. (Wikipedia; also google “Blaschko’s lines” and check out the Image results for dramatic examples of how this phenomenon can manifest in humans)
More speculation on the pied gene and multiple-gene interactions. Terry Martin has hypothesized that there are 10-20 genes involved in producing pied patterns, including major genes (both dominant and recessive) that produce significant patterns, and minor genes (again, both dominant and recessive) that produce smaller patterns, as well as modifier genes that don't directly produce a pattern but have an effect on the pattern genes.
It seems to me that all these genes probably have some kind of influence on the migration of melanoblasts or perhaps on their survival, resulting in a sort of local control where some parts of the body respond to the pied gene(s) more easily than others. The back of the head responds so easily that a pied split often causes markings there. The upper part of the wings seems to be the most resistant to the pied gene, and is the body part that is most likely to be solid grey on heavy pied birds. But this seems to be the body part that is most affected by the specific-pattern tricolor phenomenon.
Speculation on color changes over time
It seems possible that gynandromorphy might be involved in at least some cases where the bird’s color changes over time; specifically in cases where a hen experiences male-style plumage changes, or conversely in cases where a male retains female-style markings in adulthood. Since many of the examples are pied, retention of juvenile markings by males could simply be the pied mutation's typical interference with the normal sexual dimorphism. It’s also possible that a metabolic disorder or dietary deficiency was involved in the original color difference, and it got better or worse over time. Genetic factors could be involved too; there are some mutations where the coloration typically develops and changes over time.
Other info on mosaicism that probably isn’t relevant. In many mammal species, all females have a type of mosaicism caused by one X chromosome being randomly inactivated in every cell of the body; in some cells the paternal X is inactivated and in other cells it’s the maternal X. This is what causes the tabby pattern in female cats. However there is no indication that a similar phenomenon occurs in birds. (Onsman)
When all is said and done, we still don't know what causes this coloration. But even though tricolor isn't inheritable, it seems likely that there is a genetic cause in at least some cases.
*******************
(Back to Top)
 Bird #1A and 1B |
Source:
Terry
Martin’s cockatiel mutations book, page 175
Sex: Male
Known mutation genes: Possible grey pied split cinnamon
“half-sider” per Terry Martin
Parents: unknown
Breeding history: produced ordinary-colored young |

Bird #2A, age 25 days
|
Source:
Criadouro Pinheiro (English translation
here.)
Sex: Female
Known mutation genes: Whiteface pied; does not appear
to be pearl; may or may not be cinnamon.
Parents: Father is whiteface split to cinnamon, pied
and pearl. Mother is whiteface cinnamon split to pied
Breeding history: produced ordinary-colored young Note the color changes on the face and back over time; the face now looks rather similar to a male
|

Bird #3
|
Source: Criadouro Pinheiro (English translation here.) Sex: Unknown Known mutation genes: Whiteface pearl pied Parents: Unknown Breeding history: Unknown |
 Bird #4A
|
Source: ICR Facebook page Sex: Unknown Known mutation genes: Pied. Possible whiteface split. Will be split cinnamon pearl if male. Parents: Father is pied split whiteface. Mother is cinnamon pearl pied Breeding history: None (still a baby at time of posting) This chick has paler coloring in the 'shoulder' area of the wing. |
 Bird #5 |
Source: ICR Facebook page Sex: Unknown Known mutation genes: Whiteface pied Parents: Unknown Breeding history: Unknown |
 Bird #6A "Jett" before molt
|
Source: ICR Facebook page and NCS Facebook page. April Howes, owner
Sex: Male
Known mutation genes: Whiteface pied split lutino cinnamon
pearl. Visually shows both normal gray and cinnamon markings; also
seems to be showing some pearl. Any visible lutino would be
indistinguishable from pied. Possible color changes over time? Hard
to tell because the lighting and angle is so variable. |
 Bird #7A "Picasso"
|
Source:
Bert's posts on a now-closed ICR forum. More pictures are
available at these links in Bert's Photobucket account:
#1(juvenile plumage),
#2(back),
#3(left side),
#4(right side) Sex: Male Known mutation genes: Pied, apparently split whiteface.
Has some visible pearls in the cinnamon part of his plumage. But
the cinnamon and pearl genes maybe not be configured the way
one would normally expect, because the coloring of his offspring
doesn't match the expected results. Odd breeding results could be
due to a series of crossovers. Parents: Father is pied split to whiteface and to pearl
cinnamon yellowcheek on the same X; mother is pearl cinnamon pied. Breeding history: No tricolor chicks. Young inherit cinnamon OR pearl, not
both mutations together. His mate is lutino pied split to whiteface. The conversation at the end of this article
discusses this peculiar outcome extensively. |
 Bird #8A "Mickey Mouse" as a juvenile
|
Source: Susanne Russo’s picture album here and here and also the ICR Facebook page Sex: Male Known mutation genes: Creamface pied Parents: Not reported Breeding history: Not reported Note the color changes over time: apparent reduction in the medium color in the center of the back, and the addition of white along the quill in some brown wing feathers. The juvenile wing coloring is easier to see in the picture for Birds #15 below. |
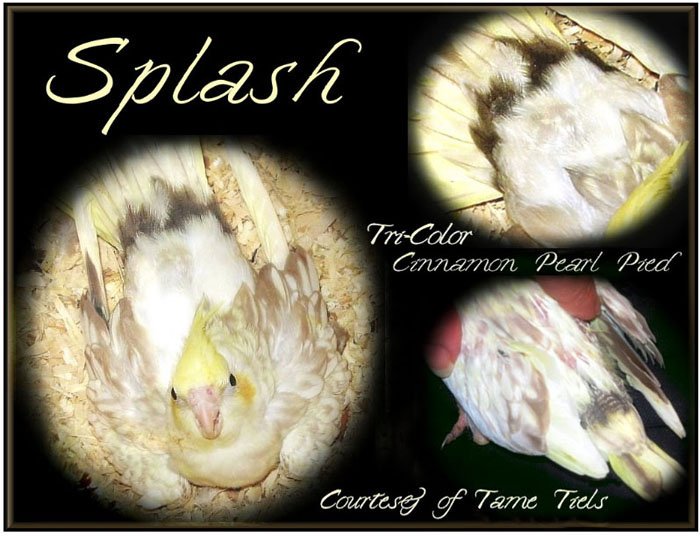 Bird #9 "Splash"
|
Source:
Susanne Russo’s cockatiel mutations e-book
Sherri Tame Tiels, owner. Sex: Not reported Known mutation genes: Not reported
Parents: Not reported
Breeding history: Not fully reported; maternal grandmother
of Bird #10 "Calico Kitty" |
 Bird #10 "Calico Kitty"
|
Source:
Susanne Russo’s cockatiel mutations e-book
Sex: Female Known mutation genes: Whiteface pied Parents: Mother is whiteface lutino, and is the daughter
of Bird #9 "Splash" Breeding history: Not reported |
 Bird #11 |
Source:
Terry Martin’s cockatiel mutations book,
page 174
Sex: Uniform face coloring on both sides indicates male Known mutation genes: Not reported Parents: Not reported Breeding history: Not reported This bird is a quartersider who can reasonably be described as tricolor.
|
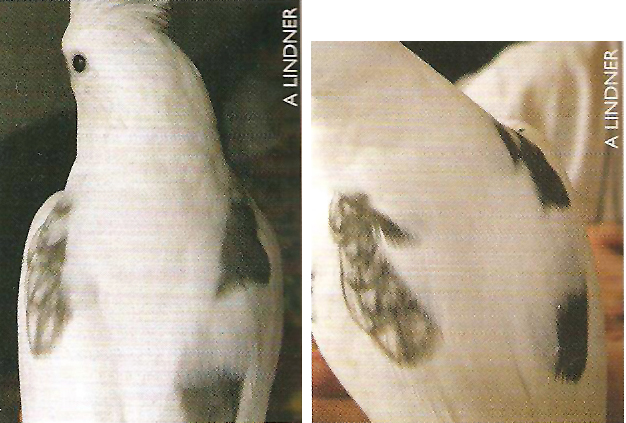 Bird #12A and B
|
Source:
Terry Martin’s cockatiel mutations book,
page 174
Sex: Unclear; might be a gynandromorph and/or
halfsider Known mutation genes: Whiteface pied with pearl markings
on on wing but not on the other Parents: Not reported Breeding history: Not reported This bird does not appear to be a tricolor. The pearled wing looks paler than the unpearled
wing simply because of the white markings on the feathers. |
 Bird #13A and B
|
Source:
Wyjątkowa nimfa with further discussion at
Genetics-Psittacine
Sex: Female
Known mutation genes: Can't be determined with any
confidence; see discussion at Genetics Psittacine link above.
Parents: Unknown
Breeding history: Lays fertile eggs. "Passes on
genotypes from both sides of her body through her gametes"
which is ambiguous but probably means that some offspring resemble
one side of the body and some resemble the other side.
This bird is a halfsider but should probably not be classified as a tricolor.
The location of the bird is unknown. The pictures appeared on
a Polish website but it doesn't sound like the webmaster owns the
bird.
Translation of the Polish website (sorry, the Google Translator did
a far from perfect job):
"I just got a picture of exceptional nymphs. It is a nymph of different colors (mutations) on each body half. This phenomenon (known in English halfsider) is quite well known in the world of birds. This looks like a parrot glued to the two separate halves. The dividing line runs through the center of the torso (when is 50/50), may also be some other aspect ratio. This parrot is a phenomenon, which is an example of mosaicism. In cultures birds of different fisheries bodies are quite common in budgies. They are also recorded in canaries, Zebra and many other species (details given Hollender 1944). Birds such it is impossible to intentionally breed (and very good), they are the result of cytological case (a bird that is the opposite of the twins), and it is not a trait inherited. This problem is not fully recognized.
"For the first time I hear about this, however, the nymph! Below pictures of females nymphs, which has sent Dr. R. Brightwell from Australia.
Clearly, the right half (pictured on our left) is yellow, while the left half is devoid of yellow dye is Caspian.""
|
 Bird #14A and B
|
Source:
Babu VT (owner) and
Talk
Cockatiels forum thread Sex: Not reported Known mutation genes: Pearl Parents: Not reported
Breeding history: None This bird has normal coloring on
one side of the face and whiteface coloring on the other. This
is an example of the halfsider phenomenon that is limited to a
fairly small area of the body. This bird should probably not be classified as a tricolor. |
 Bird(s) #15
|
Source:
Susanne Russo’s cockatiel mutations e-book
Bird at top left is same as Bird #8 Sex: Not reported Known mutation genes: Not reported Parents: Not reported Breeding history: Not reported The unusual coloring of
these birds may have different causes. The bird on the right in
particular doesn't have much in common with the other birds in the
gallery except for Birds #16 below. |
 Birds(s) #16 A and B
|
Source: Screen captures from a YouTube video. Also a discussion on the Genetics-Psittacine board Sex: Male Known mutation genes: Pied splits in at least some cases Parents: Unknown Breeding history: several of the young males fathered offspring who did not show tricolor markings In 2010 the owner reported 4 mature males and 6 young males entering their first molt, all with patches of brown feathers that are replaced by similar brown feathers when they molt. At the time of purchase they were being kept indoors but their prior history is unknown. The most striking individuals from the video are shown above. An update in the genetics board thread reported that the birds molted into normal coloration after several months on a good diet. Color changes over time have been observed in some tricolor birds who are still with their original breeder, and presumably on the same diet, so diet is not necessarily relevant. But those other birds did not molt back into normal coloration. Several theories were raised about what’s going on with the #16 flock in the Genetics-Psittacine thread, none of them conclusive and most of them not very convincing. But a similar case was mentioned where birds had mottled coloring like this and molted into normal coloring after their diet was improved. It seems likely that diet was an issue with the birds in these pictures. A genetic cause has not been ruled out, but it seems likely that any genetic cause would be related to health/nutritional issues and the coloration would be a side effect.
|
 Bird #17
|
Source:
Terry Martin’s cockatiel mutations book,
page 175
Sex: Male
Known mutation genes: Possible pearl? The tailfeathers look
grizzled and the photo quality is so poor that it's hard to tell
what's happening on the body.
It seems likely that this bird is mottled/grizzled not tricolor.
Parents: unknown
Breeding history: produced ordinary-colored young
|
 Bird #18A and B
|
Source: Terry Martin’s cockatiel mutations book, page 174
Sex: Male
Known mutation genes: Not reported
Parents: Not reported
Breeding history: Not reported This bird has a mottled pattern that changes with every molt. Not
a tricolor. |
 Bird #19 "Millie"
|
Source:
Jade Katherine Rowe (owner) and
NCS Facebook page Sex: Female Known mutation genes: Cinnamon split to pied Parents: Not reported Breeding history: Not reported This bird is NOT a
tricolor or a halfsider, at least not in the standard sense of the word. Most
cockatiels have darker color on the upper chest than they do on the
lower belly, and in some individuals the transition is abrupt.
This bird's coloring is believed to be normal. |











{kind=link}
*******************
(Back to Top)